Pochodzenie gatunków jest jednym z ciekawszych zagadnień dla hodowcy i nie tylko.
Jest wiele rodzajów motyli, które cechuje niewielka zmienność ubarwienia w obrębie
swojej gałęzi taksonomicznej. Już zwykły obserwator jest w stanie wizualnie sklasyfikować
przynależność do określonej grupy posiadającej wspólne i silne cechy. W świecie
naukowym, gatunki klasyfikuje się na podstawie analizy genetycznej, czy porównywania
aparatów kopulacyjnych. Nie są to jednak ostateczne i wystarczające metody, które
pozwalają na zbudowanie drzewa taksonomicznego. W wielu sytuacjach czynności te
wspomagane są przez aspekt zasięgu występowania gatunków. W większości przypadków
występowanie gatunków na danym terenie definitywnie potwierdza ustalone genetycznie
podobieństwo. Czasem jednak można się spotkać z ciekawymi przypadkami gatunków,
których obszar występowania nie potwierdza ich pokrewieństwa terytorialnego (A.luna
- A. artemis). Owo podobieństwo może również dotyczyć przystosowań środowiskowych
i pokarmowych, co w zasadzie już niekoniecznie musi się pokrywać z zasięgiem występowania
(A. dubernardi - G. isabelae).
Przykładów gatunków podobnych można by mnożyć. Jako jeden z ciekawszych uważam -
rodzaj Attacus. W jego obrębie sklasyfikowano kilkanaście taksonów, o bardzo podobnych
i charakterystycznych cechach. Poszczególne gatunki zostały już opisane na stronie,
a ich cechy zewnętrzne - wyszczególnione. Dlaczego wszystkie te gatunki są tak do
siebie podobne? W zasadzie pozostaje nam przyjrzenie się zasięgowi występowania
każdego z nich. Pokazuje to pierwsza mapa. Już pierwsze spojrzenie na nią sugeruje,
że powolne rozchodzenie się lądów spowodowało separację pragatunku, który zamieszkiwał
pierwotny, spory ląd. Ponieważ osobniki z rodzaju Attacus nie są wybornymi lotnikami,
to też odległość kilkudziesięciu kilometrów morskiej wody bezwzględnie separuje
rozdzielone populacje.
Problem pojawia się jednak wtedy, gdy okazuje się, że spora część wysp jest pochodzenia
wulkanicznego. Oznacza to tyle, że nie był to pierwotny ląd, który uległ rozsunięciu.
Każdy z nas zdaje sobie sprawę z tego, że kulę ziemską nawiedzały okresowe zlodowacenia.
Podczas ich trwania, czapy lodowe na biegunach ulegały rozszerzeniu, powodując spadek
poziomu mórz. Analiza terenu dna morskiego w obrębie Azji południowo-wschodniej,
Indonezji i Australii dostarczyła odpowiedzi na nurtujące nas pytania. Otóż obniżając
poziom wód oceanicznych, ustanowiono przebieg granic lądowych trwających podczas
zlodowacenia.
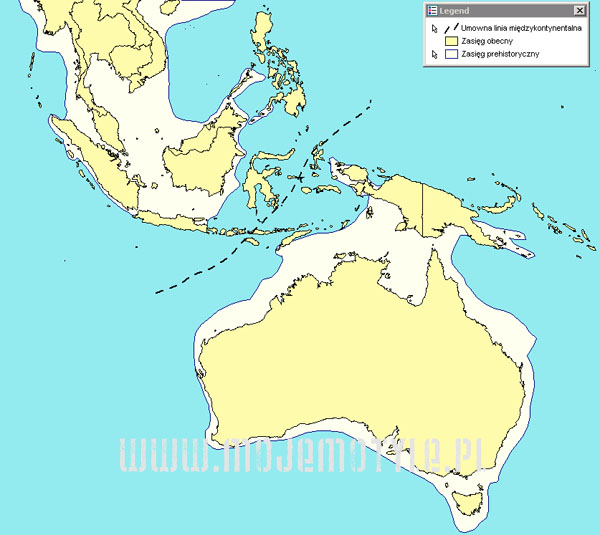
Kolejna mapa pokazuje dodatkowy ląd, który łączy już istniejące. Wyraźnie widać,
że w takim wypadku powstają nam dwie rozdzielone masy lądowe, część azjatycka i
australijska.
Nie ma sensu zagłębiać się w szczegóły i analizować, w jaki sposób i kiedy konkretny
gatunek został wyodrębniony. Jedno jest pewne, że działo się to w odpowiedniej kolejności
i zależało od odległości między wyspami. Topnienie czap lodowych spowodowało stopniowe
zalewanie obszarów nizinnych, co w konsekwencji odseparowało istniejące populacje.
Czas pełnego odosobnienia wpłynął mniej lub więcej na cechy charakterystyczne gatunków.
Oczywiście ruchy tektoniczne przyczyniły się w tym samym czasie do powstania wielu
wysp, na których owe gatunki mogły osiąść i tam się rozwijać. Natomiast większy
wpływ na ich kształtowanie miały zlodowacenia, a bezpośrednio poziom wody w oceanach.
Istnieje również kilka teorii dotyczących lini separującej faunę Indonezji od Australii
w okresie największego zlodowacenia. Na poniższej mapie przedstawiono jej uśredniony
przebieg, jednak we wspomnianych teoriach istnieje ich kilka i mają swój ustalony
przebieg i nazwę (Linia Wallace`a, Weber`a). Jednak faktyczny przebieg tej linii
jest czysto teoretycznym założeniem i nie ma on znaczenia w naszych rozważaniach.
Występowanie rodzaju attacus w Australii związane jest z jej chwilowym połączeniem
z masą lądu Nowej Gwinei. Wzrost poziomu wody odciął Australię od wilgotnego klimatu,
zostawiając jednak małe skupiska bogatej fauny na północnych krańcach kontynentu.
Tam też została odizolowana część populacji pra-Attacusa, która stała się jedyną,
pokrewną od azjatyckiego przodka.
Kolejna, ciekawa separacja nastąpiła w samej Azji. Otóż jeśli spojrzymy na mapę
występowania rodzaju Attacus, to najbardziej oddaloną populacją wydaje się być A.
taprobanis. Dlaczego tak się stało? Tutaj potrzebna jest wiedza z ruchów mas lądowych,
które kształtowały obecny rozkład mas kontynentalnych. Obszar obecnych Indii przesunął
się od południowego zachodu i zetknął się z Azją. Dzięki temu azjatycka populacja
mogła się rozprzestrzenić na nowo przybyły ląd. Niestety, masy lądu przesuwały się
dalej do przodu, powodując powolne wypiętrzanie się masywu Himalajów. Te obecnie
najwyższe góry na naszej Ziemi stały się blokadą dla mas tropikalnego powietrza
płynącego z Azji południowo-wschodniej. Wskutek tego powstał olbrzymi pas lądu nieprzyjazny
dla populacji Attacusa. Jednak dzięki nadbrzeżnemu pasowi lasów monsunowych ciągnącemu
się od prowincji Gujarat do Sri Lanki, część populacji Attacus mogła się tam utrzymać
i stworzyć odrębny gatunek.
Informacje tu przedstawione są jedynie nakreśleniem aspektu pochodzenia poszczególnych
gatunków rodzaju Attacus. Dokładna analiza wymaga nałożenia wielu czynników geologicznych,
hydrologicznych i klimatycznych na osi czasu co pozwoli na pełniejsze zrozumienie
zagadnienia. Zainteresowanych odsyłam do prac takich wybitnych naukowców jak R.
Peigler, Wallace, Diakonoff czy Mayr.